Глава 5. Происхождение океанических рыб
Общепризнано, что жизнь на Земле зародилась в прибрежной зоне океана. Там же, на мелководьях, возникли все типы и почти все классы свободноживущих животных (об этом не раз писал академик Л. А. Зенкевич), в том числе все современные классы рыбообразных и рыб - миксины, миноги, хрящевые и костные. Именно оттуда шло заселение пресных вод и суши, а также, вполне естественно, всех биотопов «открытого океана», рассматриваемых в этой книге. Меня всегда занимал вопрос, каким образом происходило это заселение. Как возникли столь существенные различия между населением донно-придонных и пелагических биотопов? Почему одни из океанических видов почти не отличаются от родственных прибрежных форм, другие хотя и имеют специфические черты, но легко выводятся от мелководных родственников, третьи относятся к аберрантным группам, положение которых в системе отнюдь не очевидно? Есть ли соответствия между степенью систематической обособленности рыб, населяющих разные биотопы открытого океана. Древностью этих рыб и очередностью заселения ими тех или иных биотопов? Нужно добавить еще, что палеонтологических свидетельств эволюции океанических рыб практически нет, и ответы на все возникающие вопросы могут быть получены только на основе косвенных доказательств.
Впервые я коснулся этих проблем в своей книге «Ихтиофауна океанской эпипелагиали», но вовсе не все, сказанное 20 лет назад, представляется мне верным сейчас, и в первую очередь это касается заселения глубоководной пелагиали, которое, как я думал тогда, шло из верхних горизонтов водной толщи.
Представления об эволюционной истории и расселении любой фауны зиждутся на изученности ее систематического состава, так как естественная система организмов должна, как известно, отражать их филогению. В отношении рыб и рыбообразных такая система, впрочем, еще не разработана, и пх классификация, даже на уровне самых высоких категорий, активно обсуждается и перестраивается в последние годы. «Классические» представления, синтезом которых была общепринятая вплоть до середины 60-х годов система академика Л. С. Берга, претерпели очень существенные изменения. В этой книге используется в основном классификация, принятая в недавней сводке Дж. Нельсона (1984) и дополненная данными более поздних исследований.(Число отрядов щележаберных, в соответствии с трудами Л. Компаньо, увеличено с 5 до 12, из отрядов карпозубообразных (Cyprmodontiformes) и бериксообразных выделены отряды сарганообразных и китовидкообразных, отряды иглообразных (Syngnathiformes), иядостомообразпых (Indostomiformes) и пегасообразных (Pegasiformes) включены в отряд колюшкообразных, а отряд долгоперообразных (Dactylopteriiormes) - п отряд снорпенообразных.)
Сопоставление систематического состава прибрежных (шельфо-неритических) и океанических таксоценов показывает значительные различия между ними. Как видно из табл. 5, в прибрежной зоне представлено 42 из 44 отрядов рыб и рыбообразных, встречающихся в соленой воде (в это число включены и такие отряды, в которых есть хотя бы отдельные диадромные виды, проводящие часть жизни в море), и только два отряда — стомиеобразные и миктофообразные в полном составе специфичны для открытого океана. В океанской пелагиали живут виды из 24 отрядов, в бентали и бентопелагиали — из 31 отряда, но далеко не все они могут считаться процветающими вдали от берегов.
Из круглоротых, например, миксины очень характерны для мезобентали, тогда как миноги (они размножаются только в пресной воде) играют ничтожную роль как в этом биотопе (трехзубая минога Entosphenus tridentatus встречается, впрочем, в северной Пацифике вплоть до глубины 1000 м), так и в эпипелагиали (Geotria australis отмечена вдали от берегов в субантарктических водах). Современные цельноголовые (кроме хоботнорылых химер рода Callorhynchus) явно тяготеют к мезобентали, хотя ископаемые формы, как полагают, были приурочены к шельфовым морям. Хорошо представлены в открытом океане щележаберные рыбы — лишь два из 12 отрядов не выходят за пределы прибрежной зоны. Заселение океанических биотопов акулами и скатами также шло главным образом вниз по склону. В мезобентали и мезобентопелагиали представлено 23 семейства из 9 отрядов, в батибентали и батибептопелагиали — 3 семейства из 3 отрядов, причем 5 семейств встречаются только на глубинах, а еще 3 имеют там центр своего разнообразия. Пелагические океанические виды существуют в 5 отрядах щележабериых (не считая гнюсообразных, случайно проникающих в открытые воды) и 9 семействах (опять же без ксеноэпипелагических); два семейства акул полностью ограничены в своем распространении эпимезопелагиалью, а одно (Lamnidae) представлено за ее пределамп только нерито-океаническим кархародоном. Из числа костных рыб в открытом океане живут виды 22 отрядов, в том числе 19 отрядов (более 140 семейств) в доннонридонпых биотопах и 17 отрядов (94 семейства) в пелагиали.
Таблица 5
| Отряды рыбообразных и рыб | Всего семейств | Прибрежные биотопы | Океанические биотопы | |||||||
| пелагиаль | бенталь и бетопелагиаль | |||||||||
| пелагиаль | пелагиаль | пелагиаль | пелагиаль | бенталь и бентопелагиаль | бенталь и бентопелагиаль | бенталь и бентопелагиаль | бенталь и бентопелагиаль | |||
| Миксинообразные (Myxiniformes) | 1 | 1 | - | - | - | - | 1 | 1 | - | - | Миногообразные (Petromyzontiformes) | 1 | 1 | 1 | - | - | - | 1 | - | - | - | Многожаберникообразные (Hexanchiformes) | 2 | 1 | - | - | - | - | 2 | - | - | - | Катранообразные (Squaliformes) | 3 | 3 | 1 | 1 | - | - | 3 | 1 | 1 | - |
| Пилоносообразные (Pristiophoriformes) | 1 | 1 | - | - | - | - | - | - | - | - |
| Плоскотелообразные (Squatiniformes) | 1 | 1 | - | - | - | - | 1 | - | - | - |
| Разнозубообразные (Heterodontiformes) | 1 | 1 | - | - | - | - | - | - | - | - |
| Воббегонгообразные (Orectolobiformes) | 7 | 7 | 1 | - | - | - | - | - | - | - |
| Ламнообразные (Lanmiformes) | 7 | 4 | 4(1) | 5 | - | - | 2 | - | - | - |
| Кархаринообразные (Carcharhiniformes) | 8 | 7 | 2 | (1) | - | - | 5 | 1 | - | - |
| Пилообразные (Pristiformes) | 1 | 1 | - | - | - | - | - | - | - | - | Гнюсообразные (Torpediniformes) | 4 | 4 | (1) | - | - | - | 2 | - | - | - | Хвостоколообразные (Myliobatiformes) | 8 | 7 | 1(1) | - | - | - | 1 | - | - | - | Скатообразные (Rajiformes) | 8 | 8 | - | - | - | - | 4 | 2 | - | - | Химерообразные (Chimaeriformes) | 3 | 2 | - | - | - | - | 2 | 1 | - | - | Целакантообразные (Coelacanthiformes) | 1 | (1) | - | - | - | - | 1 | - | - | - | Осетрообразные (Acipenseriformes) | 2 | 1 | - | - | - | - | - | - | - | - | Тарпонообразные (Elopiformes) | 4 | 4 | - | - | - | - | 1 | - | - | - | Спиношипообразные (Notacanthiformes) | 3 | (1) | - | - | - | - | 3 | 2 | 1 | - | Угреобразные (Ariguilliformes) | 19 | 10 | 1 | 5 | 4 | 1(1) | 7 | 3 | 1 | - |
| Сельдеобразные (Clupeiformes) | 4 | 3 | 1(1) | - | - | -(1) | - | - | - | - |
| Конорылообразные (Gonorhynchiformes) | 4 | 2 | - | - | - | - | - | - | - | - |
| Сомообразные (Siluriformes) | 31 | 2 | - | - | - | - | - | - | - | - |
| Карпообразные (Cypriniformes) | 6 | 1 | - | - | - | - | - | - | - | - |
| Лососеобразные (Salmoniformes) | 16 | 6 | (1) | 3 | - | - | 4 | 3 | 2 | - |
| Стомиеобразные (Stomiiformes) | 9 | - | (2) | 9 | 2 | 1 | 5 | - | - | - |
| Аулопообразные (Aulopiformes) | 12 | 4 | 1(2) | 8 | 1 | - | 4 | 2 | 2 | - |
| Миктофообразные (Myctophiformes) | 2 | - | (1) | 2 | 1 | - | 2 | - | - | - |
| Трескообразные (Gadiformes) | 8 | 6 | (1) | 3 | 1 | - | 7 | 2 | 1 | - |
| Ошибнеобразные (Ophidiiformes) | 4 | 3 | - | 1 | - | - | 4 | 3 | 3 | 1 |
| Жабообразные (Batrachiformes) | 1 | 1 | - | - | - | - | - | - | - | - |
| Удильщикообразные (Lophiiformes) | 16 | 5 | 1 | 10 | 10 | (4) | 5 | 3 | 2 | - |
| Присоскообразные (Gobiesociformes) | 1 | 1 | - | - | - | - | - | - | - | - |
| Сарганообразные (Beloniformes) | 7 | 4 | 3(1) | - | - | - | - | - | - | - |
| Атеринообразные (Atheriniformes) | 5 | 2 | - | - | - | - | - | - | - | - |
| Опахообразныё (Lampridiformes) | 10 | 1 | 5 | 9 | - | - | - | - | - | - |
| Бериксообразные (Beryciformes) | 11 | 5 | - | 2 | 1 | - | 10 | 3 | 1 | - |
| Китовидкообразные (Cetomimiformes) | 4 | (1) | - | 3 | 1 | (1) | 1 | - | - | - |
| Солнечникообразные (Zeiformes) | 6 | 2 | - | - | - | - | 7 | 1 | - | - |
| Колюшкообразные (Gasterosteiformes) | 11 | 10 | 1(1) | - | - | - | 1 | - | - | - |
| Скорпенообразные (Scorpaeniformes) | 21 | 17 | 1 | 1(1) | - | - | 13 | 4 | 1 | 1 |
| Окунеобразные (Perciformes) | 150 | 122 | 14(2) | 5(11) | 2 | - | 41 | 6 | 1 | - |
| Камбалообразные (Pleuronectiformes) | 7 | 7 | - | - | - | - | 5 | - | - | - |
| Скалозубообразные (Tetraodontiformes) | 8 | 6 | 4 | (1) | - | - | 2 | - | - | - |
| Всего отрядов | - | 42 | 21 | 17 | 9 | 4 | 31 | 14 | 9 | 2 |
| Всего отрядов | - | 277 | 57 | 81 | 23 | 8 | 147 | 38 | 16 | 2 |
В целом ихтиофауна открытого океана образует значительную по объему и хорошо обособленную часть мировой фауны рыб (табл. 6). Она включает, по самым предверительным подсчетам (для более точных оценок время еще не наступило), около 1100 видов, живущих в толще воды без какой-либо связи с дном и около 2800 видов бентопелагического комплекса (без талассосублиторальных видов, число которых, по самой грубой прикидке, составляет не менее 300—400), в том числе почти 1700 видов, не выходящих на шельф. В полном объеме океаническая ихтиофауна содержит, таким образом, около 3900 (2200 без шельфо-батиальных форм) видов (см. табл. 2—4), т. е. составляет более четверти всех морских рыб (прибрежных рыб насчитывается около 9,5 тыс. видов). Несколько отличающиеся оценочные цифры приводит Д. Коэн, по подсчету которого число видов прибрежных рыб достигает 9100, а океанических — 2500 (в том числе 255 эшшелагических, 1280 глубоководных донно-придонных и 1010 глубоководных пелагических) .
Таксономическая обособленность отдельных биотопических группировок океанических рыб довольно сильно различается. Для эпипелагиали, например, специфичны всего 16 семейств рыб (30% от числа представленных в биотопе), причем большая их часть характерна для ее нижних слоев, пограничных с мезопелагиалью. Полностью океаническим является, строго говоря, только одно из верхнеэпипелагических семейств — макрелещуковые. Специфичность на родовом и видовом уровнях достигает тем не менее 50% списочного состава.
Глубоководные пелагические ихтиоцены значительно более своеобразны. Их самобытный облик определяется главным образом наличием ряда характерных таксонов высокого ранга, таких, как отряды стомиеобразных, миктофообразных, китовидкообразных, подотряд глубоководных удильщиков и др., очень разнообразных в видовом отношении. Все они отсутствуют в прибрежной зоне и очень слабо представлены в толще воды над материковым склоном. Половина мезобатипелагических семейств (42 из 83), большинство родов (61%) и почти все виды (85%) также специфичны для этих биотопов.
Придонные ихтиоцены открытого океана представляются менее обособленными, но и они имеют характерные отряды (целакантообразные, спиношипообразные) и подотряды (например, гладкоголововидные). Число специфических семейств относительно невелико — всего 20 % (27 из 137), хотя на уровне родов и видов эти таксоцены вполне четко отличаются от прибрежных и пелагических, а также друг от друга.
В целом наиболее резко выражены границы между совокупностями пелагических и донных таксоценов рыб, причем степень выраженности этих границ явно увеличивается с глубиной. В вертикальной последовательности бентальных и бентопелагических ихтиоценов самая четкая разделительная полоса соответствует краю континентальной отмели, но, как указывалось в предыдущей главе, многие преимущественно шельфовые виды все же опускаются в верхние участки материкового склона. Дальнейшие изменения таксономического состава — от мезобентали к батибентали и далее к абиссобентали и хадали — происходят постепенно и наиболее заметны на видовом уровне. В толще воды границы между эпи-, мезо- и батипелагиалью, несмотря на существование зон смешения (экотонов), выражены более четко, чем границы между гомологичными донно-придонными биотопами.
Исходным материалом для формирования рецентной фауны океанических рыб бесспорно были шельфовые и неритические предковые формы. Из этого постулата со всей определенностью следует, что именно прибрежные участки (или по меньшей мере некоторые из них) действовали в определенные периоды времени, а быть может, действуют и сейчас как эволюционные центры, продуцирующие виды, способные к заселению новых биотопов за пределами этих участков, т. е. к переходу в новую среду обитания. (Альтернативой такому представлению могло бы служить только явно абсурдное в данном аспекте предположение о некогда сплошном распространении предковых форм в относительно однородной среде и последующем расчленении их ареалов в результате образования физических барьеров (так называемая «викариация» в понимании сторонников кладизма),) Очевидно также, что прямо из шельфо-неритической зоны могли заселяться только непосредственно прилегающие к ней биотопы, которые, в свою очередь, обеспечивали заселение контактирующих с ними участков среды. Последовательность появления вселенцев в том или ином биотопе может в первом приближении определяться их ранжированием по систематической обособленности: наличие видов, общих с исходным (донорским) биотопом, говорит о совсем недавнем или еще не закончившемся вселении, тогда как дивергенция на видовом и родовом уровне, а тем более на уровне семейств, подотрядов и отрядов свидетельствует о все увеличивающемся сроке, прошедшем со времени успешной колонизации.
Основываясь на этих представлениях, рассмотрим теперь, как происходило заселение донно-придонных океанических биотопов. В мезобентали очень обычны рыбы, характерные и для шельфовой зоны (см. табл. 4). Большинство этих эврибатных эпимезопелагических видов входит в состав экотонного сообщества кромки шельфа и самой верхней батиали и не опускается глубже 400— 500 м. Среди них, несомненно, есть и такие формы, которые потенциально способны к адаптивной модификации, дающей возможность закрепиться на глубинах материкового склона.
Примеры такого закрепления показывают мезобентальные виды преимущественно шельфовых родов, например Saurida kaianus (сем. Synodontidae) или Chaetodon guyotensis (Chaetodontidae). Есть в этом биотопе и специфические роды, принадлежащие к преимущественно шельфовым семействам — Ostichthys (сем. Holocentridae), Aphanopus и Assurger (Trichiuridae), Bembradium (Platycephalidae) и др. К эндемичным мезобентальным семействам относятся плащеносные акулы и акулы-домовые, ложноромбовые и шестижаберные скаты из щележаберных, глубоководные солнечники, хаунаксовые и хол-лардиевые из костистых рыб. Только в мезобентали сохранился до нашего времени и единственный представитель кистеперых рыб — латимерия.
В глубине души я всегда надеялся увидеть латимерию живьем, но мне так и не довелось, к превеликому сожалению, присутствовать при ее поимке. Ближе всего к живому целаканту я был, пожалуй, в апреле 1983 г., когда судно «Рифт» проводило траления на глубине 400-500 м у северо-западного побережья Мадагаскара, всего в какой-нибудь сотне миль от Коморских островов. Увы, безрезультатно... Правда, с фиксированным экземпляром мне познакомиться все-таки довелось. Дело в том, что еще в 1972 г. наш институт приобрел по специальному указанию директора — члена-корреспондента Академии наук А. С. Монина, одну латимерию для предполагавшейся выставочной коллекции. Покупка была оплачена через Париж (острова еще были колонией Франции), но для получения экспоната «Дмитрий Менделеев», работавший тогда в Индийском океане, был послан прямо на остров Гранд-Комор. Вот как описывает этот эпизод участник экспедиции и мой хороший друг Виктор Нейман: «Разрешения от местных властей на посещение Коморских островов мы в тот раз не получили и должны были принять рыбу на борт на внешнем рейде... После довольно долгого томительного ожидания к судну подошел, наконец, старенький катер, на палубе которого в открытом жестяном ящике находилась с нетерпением ожидавшаяся всеми диковина. Вид ее нас, однако, разочаровал до крайности - грязно-рыжая рыбина, по-видимому, давным-давно ждавшая нас в растворе формалина, отнюдь не блистала красотой». Действительно, целакант прибыл в Москву в ужасном состоянии — его полутораметровое тело было почти полностью покрыто плотной, как панцирь, коркой красноватой ржавчины, удалить которую механическим путем, не повредив при этом внешних покровов, было невозможно. Очистку латимерии по предложению сотрудника нашей лаборатории Дмитрия Астахова, по праву заслужившею звание «лучшего химика
среди ихтиологов», вели с применением растворителя окиси железа — так называемого трилопа Б. С результатами этой «реставрационной деятельности» можно ознакомиться в фойе Института океанологии им. П. П. Ширшова (Москва, ул. Красикова, 23), где обновленный целакант в гордом одиночестве выставлен для обозрения в большом плексигласовом сосуде, доверху заполненном фиксатором. Жаль только, что рыба навсегда утратила свою сирепево-синюю окраску и стала теперь тускло-серой, но такова уж судьба всех музейных экспонатов.
Существование особых мезобентальных семейств, мезобентальных родов в шельфо-батиальных семействах и мезобентальных видов в преимущественно шельфовых родах, а также проникновение отдельных шельфовых видов в верхние участки материкового склона свидетельствует о большой растянутости (в историческом аспекте) процесса заселения глубин. Как правило, мезобеитальные таксоны принадлежат к числу наиболее генерализованных представителей своих групп (отряд Hexanchiformes в подклассе щележаберных, семейство Synaphobranchidae в отряде угреобразных, семейство Moridae в отряде трескообразных, семейство Triacanthodidae в отряде скалозубообразных, семейство Chaunacidae в отряде удильщикообразных, род Aphanopus в семействе рыб-сабель), хотя имеются и обратные примеры (например, все батиальные роды колючих акул более специализированы, чем преимущественно шельфовый род Squalus). По мнению Дж. Бриггса, заселение мезобентали происходило главным образом в результате вытеснения с шельфа примитивных форм, не выдержавших конкуренции с новыми более высокоорганизованными рыбами, и в последующем сопровождалось видовой радиацией этих вселенцев на новой родине. С этим заключением вполне можно согласиться. Нужно отметить, однако, что в толще воды над верхней частью склона, помимо рыб, происходящих от неритических предков, очень существенную роль играют макропланктонные виды океанического происхождения (миктофиды и др.), о которых будет сказано ниже.
В биотопах нижней части склона таксономическое разнообразие рыб значительно меньше, чем в верхней. Это касается как видов, так и семейств, а на абиссальных глубинах их число еще ниже (см. табл. 6). Еще более обеднен хадальный ихтиоцен, в котором представлены всего 8 видов из двух семейств. Систематическая обособленность всех вертикальных группировок донно-придонных глубоководных рыб сравнительно невелика, и их происхождение не вызывает сомнений. Батибентальная ихтиофауна сформировалась из мезобентальной в результате опускания предковых форм вдоль материкового склона, абиссобентальная ведет свое происхождение от батибентальной, а хадальная — от абиссобентальной. На всем этом пути практически не образовывалось новых таксонов выше родового уровня — в батибентали и более глубоких биотопах нет ни одного специфического семейства рыб (иногда относимые к таковым моногнатовые, афионовые и стефанобериксовые содержат и мезобентальные виды) и даже специфические роды очень редки, что позволяет предполагать относительную молодость всей этой фауны. Ихтиоцены больших глубин сформированы только костистыми рыбами (самые глубоководные хрящевые рыбы из числа колючих и кошачьих акул, ромбовых скатов и носатых химер редки в нижней части склона и вовсе не опускаются на ложе океана), принадлежащими к весьма ограниченному контингенту семейств. В абиссобентали и бентопелагиали, например, представлено всего 13 семейств, из которых наибольшим числом видов выделяются два — ошибневые (около 45) и долгохвостовые (35), и оба они распространены в чрезвычайно широком диапазоне глубин.
Рассматривая вертикальное распределение батиальных и абиссальных рыб, легко заметить, что в заселении глубин Мирового океана приняли участие отряды и семейства, не находящиеся между собой в близком родстве и колонизировавшие новые для них биотопы независимо друг от друга. Формирование ихтиофауны океанских глубин происходило, судя по географическому распространению и экологическим предпочтениям родственных мелководных таксонов, за счет двух четко обособленных источников — тепловодно-тропического и холодноводного. Из числа обитающих в бати- и абиссобентали рыб явно имеют тропическое происхождение представители отрядов кархаринообразных (кошачьи акулы Apristurus), спиношипообразных, угреобразных, аулопообразных, ошибнеобразных, удильщикообразных, бериксообразных и солнечникообразных, которые и на шельфе не представлены за пределами тропической зоны; также от тепловодных предков, по всей вероятности, ведут свое начало глубоководные химеры (отряд химерообразные), колючие акулы (отряд катранообразные) и долгохвосты (отряд трескообразные). С другой стороны, не вызывает сомнений субполярное происхождение глубинных ромбовых скатов (отряд скатообразные) и колючеперых костистых рыб — липарид, психролютид (отряд скорпенообразные) и зоарцид (отряд окунеобразные), все мелководные родичи которых обитают сейчас на шельфах холодных морей. Не столь ясно положение с гладкоголовыми рыбами (сем. Alepocephalidae), которые принадлежат к древнему отряду лососеобразных, ныне представленному на мелководьях только в умеренных водах обоих полушарий, но, возможно, имевшему в прошлом сплошное распространение в прибрежной зоне. Гладкоголовов вместе с двумя близкими семействами обычно выделяют в особый подотряд Alepocephaloidei, бесспорно родственный мезопелагическим аргентиновидным рыбам (подотряд Argentinoidei) — иногда эти подотряды даже объединяют в один, имевший с ними общих бесследно вымерших предков. Каков был облик этих мелководных предков, где они обитали и как вселялись на глубины, можно только догадываться (мне кажется, что их следует все же поискать среди ископаемых меловых или нижнепалеогеновых «сельдеобразных», населявших теплые воды). Основные центры формирования глубоководной ихтиофауны находились, как можно допустить по аналогии с другими группами животных, во-первых, в районе современного Индо-Малайского архипелага и, во-вторых, в северной Пацифике, тогда как антарктический эволюционный центр, игравший, как полагают, существенную роль в становлении глубинной фауны бентосных беспозвоночных, не был столь же действенным в отношении донных рыб.
Современная эпипелагическая ихтиофауна ведет свое происхождение от прибрежных, главным образом неритическо-пелагических, предковых форм, хотя в ее составе имеются также отдельные роды и виды, происходящие из преимущественно донных и придонных групп (например, скат-хвостокол Dasyatis violacea, острохвостый угорь Веnthenchelys cartieri, рыба-клоун Histrio histrio). Все специфические голоэпипелагические семейства костистых рыб принадлежат к отрядам сарганообразных, опахообразных, окунеобразных и скалозубообразных. Каждый из них имеет в своем составе и неритические семейства, как правило, более генерализованные по сравнению с океаническими. Голоэпипелагические роды в широко распространенных семействах также, как правило, принадлежат к числу их наиболее специализированных представителей: в качестве примера можно назвать летучих полурылов Oxyporhampbus в сем. полурыловых, летучек Exocoetus и Hirundichthys в сем. летучих рыб, рыб-лоцманов Naucrates в сем. ставридовых, тунцов Allothunnus и Thunnus в сем. скумбриевых (самый примитивный род этого семейства — Gasterochisma тоже океанический, но распространен в южном полушарии на крайней периферии ареала группы, что вполне объяснимо его вытеснением из тепловодной зоны вышеорганизованными формами), роды Centrolophus и Icichthys в сем. центролофовых. В то же время голоэпипелагические виды, входящие в состав широко распространенных родов Сагcharhinusv Dasyatis, Stolephorus, Cheilopogon, Canthidermis, Lagocephalus, Diodon, сравнительно слабо отличаются от своих неритических родственников по уровню специализации.
Различия в уровне таксономической обособленности современных голоэпипелагических рыб и прибрежных родственных им форм показывают, что колонизация океанской эпипелагиали представляет собой перманентный, очень растянутый во времени процесс, продолжающийся, судя по встречаемости в открытом океане видов, общих с неритической зоной, до настоящего времени. В этом случае мезоэпипелагические и ксеноэпипеелагические рыбы должны рассматриваться в качестве очередных «разведывательных групп», ведущих освоение открытого океана.
Трудно, пожалуй, вообразить себе рыбу, менее соответствующую классическому представлению об обитателе эпипелагиали, чем диодоны, или рыбы-ежи, из рода двузубов. Все они очень похожи друг па друга. Их короткое, на первый взгляд довольно неуклюжее тело покрыто длинными острыми колючками, которые плотно прижимаются к коже при спокойном плавании. Диодоны способны раздуваться до шарообразной формы, наполняя воздухом или водой особый мешок (вырост желудка), а их колючки при этом встают торчком, как у настоящих ежей, и превращаются в эффективное оружие пассивной защиты. Двузубы - типичные обитатели прибрежных мелководий; их зубы преобразованы в сплошные твердые пластины (по одной в каждой челюсти — отсюда и название этих рыб), очень полезные для разгрызания иглокожих, моллюсков, кораллов. В то же время мальки некоторых рыб-ежей довольно обычны в эпипелагиали, а один вид - океанический двузуб (Diodon eydouxii), внешне мало отличающийся от других сородичей, постоянно живет в открытом океане, и мощные зубные пластины нисколько не мешают ему питаться мелким планктоном. Диодонов часто удается наблюдать на световых станциях, и, должен сказать, в воде эти рыбы отнюдь не выглядят очень уж неуклюжими. Они с большим проворством пересекают освещенную зону и довольно ловко уклоняются от закидного сачка, но даже легкое прикосновение к ежу вызывает у него совершенно неадекватную реакцию: он немедленно превращается в колючий шар, который, может быть, трудно проглотить хищнику, но очень просто достать из воды любой сеткой.
На мой взгляд, океанический двузуб самим фактом своего существования представляет одно из наиболее наглядных доказательств продолжающегося заселения эпипелагиали прибрежными рыбами. Это — пример недавнего вселенца, нашедшего в новом биотопе благоприятные условия, но еще не успевшего приобрести специфических адаптации к нему. На противоположном полюсе я поместил бы луваря (Luvarus imperialis) (рис. 43) — удивительную, внешне ни на кого не похожую рыбу, которую в продолжение многих лет считали родственной скумбриевым или ставридовым. Новейшие анатомические исследования показали, что луварь ведет начало от прибрежных рыб-хирургов, но длительная эволюция в условиях открытого океана сделала его совсем непохожим па своих предков.
Рис. 43. Луварь - рыба, родственные связи которой долго оставались непонятными.
В эпипелагиали явно преобладают филогенетически молодые группы костистых рыб (в частности, окунеобразные). Основываясь на этом факте, я предполагал ранее, что до появления этих вселенцев верхние слои открытого океана были заселены низшими (мягкоперыми) рыбами, которые, не выдержав конкуренции, оказались вытесненными вниз и дали начало современному населению средне-глубинной зоны. Сейчас я считаю это умозрительное предположение крайне маловероятным. Существует гораздо больше оснований для иной точки зрения: главным источником формирования мезопелагического ихтиоцена послужили придонные и придонно-пелагические формы, обитавшие на материковом склоне.
Представление о происхождении мезопелагических рыб от мезобентальных и мезобентопелагических предков основывается на следующих соображениях. Специфические подотряды и семейства мезопелагических рыб относятся к таким отрядам (угреобразные, аулопообразные, миктофообразные, трескообразные, бериксообразпые и др.), основная масса представителей которых обитает на континентальном склоне, и являются наиболее специализированными таксонами этих групп. Семейство Chiasmodontidae наиболее родственно верхнемезобенталыюму сем. Champsodontidae. Мезопелагические роды в семействах, распространенных и вне рассматриваемого биотопа, принадлежат к группам, характерным в основном для мезобеитали (Isistius и Euprotomicrus — к сем. колючих акул, Хеnорhthalmichthys — к сем. серебрянковых, Scopelengys — к сем. неоскопеловых, Brotulotaenia — к сем. ошибневых, Супо-macrurus и Mesobius — к сем. долгохвостовых, Diretmus — к сем. диретмовых, Diplospinus и Nealotus — к сем. гемпиловых, Ectreposebastes — к сем. скорпеновых, Nectoliparis и Rhodichthys — к сем. морских слизней). В то же время мезопелагические виды в преимущественно придонных родах, по-видимому, очень редки: единственный известный мне пример представляет Nezumia parini, — описанный К. Хаббсом и Т. Ивамото пелагический долгохвост из восточной тропической Пацифики.
Бати- и абиссопелагический ихтиоцены в основном формировались, по-видимому, тем же путем, что и мезопелагический. Судя по распространению рецентных родственных групп, специфические высокоспециализированные глубоководные семейства угреобразных (пилосошниковые, циемовые, мешкоротовые) и удильщикообразных (11 семейств подотряда Ceratioidei) ведут свое начало от донно-придонных рыб нижней части материкового склона. В пелагиали больших глубин значительная роль принадлежит и представителям таких семейств, которые еще не полностью утратили связь с дном — гладкоголовам и платитроктовым, откладывающим донную икру, а также долгохвостам, афионидам и др. В то же время наличие в составе глубинной пелагической ихтиофауны родов, принадлежащих к преимущественно мезопелагическим семействам гоностомовых, малакостеевых и миктофовых, указывает на частичное заселение этой зоны из средних горизонтов.
Нужно сказать, что представители отдельных групп мезопелагических рыб, принадлежащих к макропланктону, оказались способными выйти за пределы исходного биотона и значительно расширить адаптивную зону обитания этих групп. Наиболее характерные примеры внедрения во все смежные участки среды дают светящиеся анчоусы (сем. миктофовые) которые справедливо считаются одной из самых процветающих групп мезопелагического ихтиоцена. Тем не менее около трети видов семейства (а их число достигает 220) временно или постоянно входит в состав других сообществ. Так, около 40 видов поднимаются ночью в эпипелагиаль (по меньшей мере один из них — Benthosema pterotum, наблюдавшийся в восточной тропической Пацифике у поверхности даже в светлое время суток, может образовывать там жилую, т. е. не мигрирующую, популяцию), около 10 видов живут в батипелагиали и около 20 — вторично возвратились в бентопелагические биотопы (туда, где когда-то обитали предковые формы миктофид) и в некоторых случаях достигли там высокой численности. Вторичнобентопелагические виды возникли и в других мезопелагических семействах — среди топориковых (достаточно вспомнить мавролика), фотихтовых, гоностомовых, меланостомиевых, меламфаевых. Такие виды есть также среди мезобатипелагических, в основной своей массе глубоководных удильщиков церациоидей: это представители родов Diceratias и Phrynichthys. Мало того, один из видов сем. тауматихтовых — Thaumatichthys axeli является, по всей вероятности, вторичнодонной рыбой (единственный в своем роде случай «бентизации», контрастирующий с обычной «пелагизацией» генеративно донных форм).
Говоря о заселении рыбами океанических биотопов, нельзя обойти вниманием вопрос о параллелизмах и конвергенциях в их эволюции, сопровождавших освоение тех или иных участков среды обитания. Этот вопрос, издавна привлекавший внимание исследователей, был в недавнее время детально разобран Н. Маршаллом, что позволяет ограничиться здесь рассмотрением отдельных, наиболее ярких проявлений функционального и структурного сходства систем и органов у рыб, входящих в состав разных сообществ. В эпипелагиали наиболее поражает разительное сходство между акулами ламнидами (к ним относятся, напомню, сельдевые акулы, мако и кархародон) и тунцами. Это касается не только общей формы идеально обтекаемого торпедообразного тела, его окраски, положения и относительных размеров непарных плавников, наличия стабилизирующих килей на чрезвычайно узком хвостовом стебле, несущем мощный хвостовой плавник в форме полумесяца. Еще более замечательны конвергенции в строении внутренних органов, и главная из них несомненно состоит в необычайном развитии подкожной сосудистой системы, питающей туловищную мускулатуру и обеспечивающей «теплокровность» у этих совсем не родственных рыб. И у тунцов и у ламповых акул мощно развиты наиболее активные красные мышцы латеральной мускульной системы. Волокна этих мышц окружены «чудесной сетью» капилляров, которая обеспечивает интенсивный теплообмен между артериальной кровью, исходно (в жабрах) имеющей температуру морской воды, и венозной кровью, нагретой за счет мышечной работы. В результате этого обмена температура тела тунцов и акул оказывается па 6—10° выше «забортной» температуры, и это приводит к значительным энергетическим выгодам (ведь скорость всех ферментативных реакций повышается в таком случае в 2—3 раза). Крупные тунцы (виды рода Thunnus) и акулы сем. Lamnidae могут считаться «идеальными нектерами» — как по «крейсерским» достоинствам, так и по максимальной скорости на спринтерской дистанции они не имеют соперников ни среди других рыб (исключение составляют мечевидные, достигшие тех же результатов), ни среди кальмаров и морских млекопитающих.
Примеры конвергенции и параллелизмов среди глубоководных пелагических рыб очень многочисленны, и их полное перечисление заняло бы здесь слишком много места. Нельзя не упомянуть тем не менее бросающегося в глаза сходства в окраске тела — серебристого у многих верхыемезопелагических видов (из батилаговых, топориковых, миктофовых, палочкохвостых и др.) и черного или коричневого у всех форм, обитающих на глубине более 1000 м. Органы свечения независимо возникли у колючих акул (отряд катраыообразные) и у костистых рыб, принадлежащих к отрядам лососеобразных, стомиеобразных, аулопообразных, миктофообразных, окунеобразных (род Pseudoscopelus). Телескопические глаза характерны для представителей отрядов лососеобразных (Xenophthalmichthys, Opisthoproctus), аулопообразных (Gigantura, Scopelarchus, Evermannella), миктофообразных (Protomyctophum), опахообразных (Stylephorus). Многие стомиеобразные и все удильщики приманивают добычу своеобразными удочками со светящейся приманкой. Для всех хищников характерны приспособления для захвата крупной добычи — огромные пасти, зубатые челюсти, редуцированные жаберные тычинки, сильно растяжимые желудки и стенки тела. У идиакантов и удильщиков карликовые самцы значительно мельче самок. Параллельным путем возникли и вертикальные миграции мезопелагических рыб.
Многие рыбы, обитающие в придонных слоях (галозавры, долгохвосты, ателеопы, ошибневые), независимо приобрели очень сходный тип строения, для которого характерны выступающее рыло, нижний рот, очень длинная, постепенно сходящая на нет хвостовая часть туловища, несущая по нижнему краю многолучевой аномальный плавник. Ундуляции такого плавника переводят рыбу в наклонное (головой вниз) положение, очень удобное для поиска корма на грунте. Все эти рыбы обладают плавучестью, близкой к нейтральной, как, впрочем, и ведущие сходный образ жизни придонные колючие акулы, но конвергентно сходный результат достигается принципиально разными путями — все перечисленные костистые рыбы имеют плавательный пузырь, а хрящевые регулируют плавучесть высоким содержанием легкого жира в огромной печени.
Скудность палеонтологического материала, значительные в ряде случаев отличия океанических таксонов рыб от родственных прибрежных форм, а также их сходство между собой, определяемое конвергенциями и параллелизмами, создают определенные трудности для воссоздания эволюционной истории рыб открытого океана. Тем не менее происхождение рецептной океанической ихтиофауны вполне очевидно: исходным материалом для ее формирования были шельфовые и неритические предковые формы. Заселение придонных глубин и открытой пелагиали, осуществлявшееся многими группами независимо друг от друга, представляется очень растянутым во времени. Датировка начала этого процесса, теряющаяся в далеком прошлом, невозможна без малооправданных спекуляций, а на примере эпипелагиали и мезобентали видно, что заселение основных океанских биотопов продолжается и поныне. В результате для каждого из этих биотопов характерна разновозрастная фауна.
Как известно, степень морфо-экологической специализации к условиям глубин, косвенно отражающая филогенетический возраст таксонов, была положена А. П. Андрияшевым в основу разделения глубоководных рыб на две группировки — древнеглубоководную и вторичноглубоко-водную. Такое разделение, несомненно реальное и очень наглядное в некоторых крайних случаях, все же довольно относительно и не дает возможности альтернативно определить принадлежность любой и каждой систематической группы рыб к одной из этих категорий. Представляется, что наиболее резко обособлены от ближайших родственников таксоны, возникшие в результате пелагизации, т. е. при переходе от бентического образа жизни к бентопелагическому, а затем к обитанию в безбрежной толще вод. Эти переходы носившие прогрессивный характер, сопровождались повышением общего уровня организации и соответствовали, таким образом, введенному академиком А. Н. Северцовым понятию «ароморфоз», тогда как постепенные адаптивные преобразования при увеличении глубины обитания не выходили за рамки частных приспособлений.
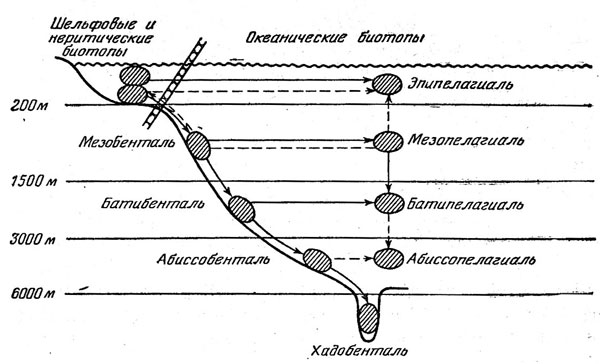
Рис. 44. Пути заселения океанических биотопов рыбами.
Генеральная схема заселения океанических биотопов представляется сейчас в следующем виде (рис. 44). Шельфовые предки дали начало мезобентальным рыбам, а от них последовательно произошли бати- и абиссобентальные и хадальные формы. В этом процессе очень существенная роль принадлежала вытеснению на большие глубины примитивных представителей тех или иных таксонов, не выдержавших конкуренции с более прогрессивными формами. Заселение разных горизонтов пелагиали происходило от придонных и неритических предков независимо на разных глубинах и шло главным образом по пути прогрессивной ароморфной эволюции; в формировании батипелагической фауны существенное значение имело и заселение сверху, из мезопелагиали. Картина современного биотопического распределения океанических рыб осложняется вторичным подъемом представителей некоторых групп в более поверхностные горизонты, вторичным же возвращением из открытой пелагиали к склонам.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://fish.geoman.ru/ 'Иллюстрированная энциклопедия рыб'