Глава 1. Открытый океан как среда обитания рыб
Представление о живом покрове Земли как об иерархичекой системе биоценозов (сообществ) дает возможность эассматривать открытый океан и в качестве единого крупномасштабного биотопа, занимающего значительную часть планеты, и в качестве системы соподчиненных биотопов все меньшего и меньшего пространственного масштаба. Расчленение океана на естественные биотопические участки определяется, как известно, тремя типами зональности — вертикальной, обусловленной неоднородностью толщи воды и изменением с глубиной подводного рельефа, широтной, в основе которой лежат климатические (в первую очередь температурные) различия, и циркумконтинентальной, обусловленной разной удаленностью от крупных массивов суши. Естественные участки океана образуют закономерно построенные биотопические комплексы, которые расположены по единому плану, повторяющемуся во всех океанах — Тихом, Атлантическом и Индийском.
В подводном строении дна выделяются три основные формы рельефа, имеющие планетарный масштаб,— материковая отмель, материковый склон и ложе океана со срединно-океаническими хребтами. Материковая (или континентальная) отмель, называемая также шельфом, представляет собой подводное продолжение материков и характеризуется очень пологим, почти незаметным увеличением глубины. За ее внешний край принимают линию, на которой угол наклона дна резко увеличивается: этот перелом кривой рельефа дна может находиться на глубине от 100 до 400 м, а его среднее положение примерно соответствует положению изобаты 130 м. Ширина материковой отмели варьирует в очень широких пределах, но составляет в среднем всего 70 км, ее суммарная площадь (до глубины 200 м) не превышает 7,5% акватории Мирового океана.
Подводный цоколь материков — материковый (или континентальный) склон имеет значительно большую крутизну. Угол его наклона в среднем составляет 3—4° (это в 25 раз больше, чем средний наклон шельфа), а местами, например у берегов Флориды, достигает даже 45°. Поверхность склона почти всегда расчленена на уступы и ступени, рассечена поперечными и косыми разломами, грядами и каньонами. Его нижняя граница обозначена новым переломом батиграфической кривой на глубине порядка 2500—3500 м (обычно около 3000 м) и соответствует переходу к ложу океана. Ширина материкового склона невелика (как правило, от 15 до 30 км), его площадь в границах между изобатами 200 и 3000 м составляет около 17,5% (без морей 12%) Мирового океана. На границе между окраинами материков и океаном в тектонически активных районах располагаются островные дуги и глубоководные желоба — очень глубокие (в Марианском желобе находится максимальная отметка — 11 022 м) и очень узкие (ширина по дну десятки километров, а по верхним частям склонов до 200 км) долины, занимающие всего лишь 0,1% площади океанов.
На ложе Мирового океана приходится около 75% поверхности его дна. Горными хребтами, возвышенностями, и валами оно разделяется на отдельные котловины — холмистые или плоские равнины, покрытые толстым слоем мягких осадков. Что касается подводных поднятий, наиболее масштабными из них являются, конечно, грандиозные срединно-океаиические хребты, простирающиеся в общей сложности более чем на 60 тыс. км. Эти хребты, центральные долины которых представляют собой границы гигантских литосферных плит, располагаются как правило, в осевых частях океанов и образуют единую систему массивных горпых сооружений. Они расчленены на отдельные блоки, сдвинутые относительно продольной оси хребта, поперечными трансформными разломами, достигающими значительной глубины. Среди других категорий поднятий следует назвать вулканические и глыбовые хребты, многие из которых имеют очень большую протяженность и высоту. Хребты всех типов, как и на суше, увенчаны вершинами и пиками. Одни из них возвышаются островами над поверхностью океана, а другие не доходят до нее. Кроме того, во всех океанах есть также изолированные горы (в основном вулканы), не связанные с массивами. Общее число подводных гор очень велико (по разным оценкам, от 8 до 10 тыс.), причем более чем у 2,5 тыс. из ниx вершины находятся на глубине менее 2000 м. Крутые склоны гор почти всегда имеют резкий, сильно расчлененный рельеф и лишены осадочного чехла, но некоторые — их называют гайотами — обладают совершенно плоскими вершинами, часто покрытыми шапкой отложений и достигающими диаметра 30—40 км. Когда-то они были островами или поднимались до уровня, на котором сказывается влияние прибоя (волновой абразии). О происшедшем погружении гайотов часто свидетельствуют и остатки коралловых построек на их вершинах и береговые террасы, выбитые прибойиымы волнами.
Особого упоминания заслуживает такая категория поднятия дна, как подводные «микрокоптиненты», отличающиеся значительными площадями малых (порядка шельфовых) глубин. Они особенно характерны для Индийскоко океана (плато Мадагаскара, Сейшельская банка, банка Сая-де-Малья); в Тихом океане к этой категории относится Новозеландское плато. По геологическому строению они существенно отличаются от других возвышенностей континентальным типом коры (с «гранитным» слоем независимо от наличия или отсутствия больших массивов суши). Объяснение существованию микроконтинентов следует искать в истории соответствующих районов Мирового океана.
Вопросы происхождения и развития океана находят объяснение с позиций глобальной тектоники литосферных плит. Раздвигание (спредипг) этих гигантских жестких блоков земной коры в обе стороны от рифтовых долин, вытянувшихся вдоль срединно-океанических хребтов, приводит к образованию новых участков дна, заполняющихся материалом мантии (движение плит происходит со скоростью до 10 см в год). При раздвижении коры образуются рифтовые впадины с рвами, расщелинами, кратерами типа «черных курильщиков», через которые на поверхность Дна изливается сульфидный гидротермальный раствор, обеспечивающий развитие специфической бактериальной флоры — хемотрофных серобактерий. Противоположной зоне спрединга край плиты давит на соседнюю плиту и, погружаясь под континент, вновь преобразуется в мантийное вещество. На основе теории тектоники плит были осуществлены палеогеографические реконструкции конфигурации и положения материков и океанов, показавшие кардинальные изменения среды существования в ходе геологической истории Земли.
Толща воды может быть условно разделена па отдельные объемы (водные массы), различающиеся по температуре, солености, плотности и многим другим физическим и химическим характеристикам. Границы между водными массами выражены с разной степенью четкости и иногда почти незаметны. В пределах водных масс и между ними постоянно происходят перемещения воды различного направления и скорости — от хаотических турбулентных движений до направленных постоянных течений. Турбулентное перемешивание толщи вод стремится размыть все гидрологические градиенты и выровнять все характеристики.
Постоянные течения переносят значительные объемы воды и в своей совокупности образуют крупномасштабные круговороты, охватывающие огромные акватории.
Характеристики водных масс формируются в система циркуляции вод, и в каждом крупномасштабном круговороте в верхнем слое переносится вполне определенная водная масса, обладающая значительной временной стабильностью и пространственной однородностью. Так, в Тихом океане Субарктическому круговороту соответствует субарктическая водная масса, Северному субтропическому круговороту — северная центральная водная масса, тропическим круговоротам — экваториальная водная масса и т. д. Такие водные массы, перемещающиеся в пределах замкнутой циркуляции и сохраняющие сравнительную однородность на больших пространствах, носят название первичных. В то же время на границах между двумя соприкасающимися круговоротами происходит смешение контактирующих водных масс и в результате образуется новая — вторичная водная масса. Этим путем формируется, например, водная масса Северо-Тихоокеанского течения. Для вторичных водных масс характерно постоянное обновление в зоне смешения и постепенная трансформация по ходу зонального течения, приводящая в конце концов к существенному изменению их характеристик. Таким образом, вторичные водные массы существуют на границах основных круговоротов и не связаны с замкнутыми циркуляциями.
В Атлантическом и Тихом океанах имеется по шесть крупномасштабных круговоротов, а в Индийском, северная часть которого подвержена существенным сезонным изменениям в связи со сменой муссонов,— четыре круговорота зимой северного полушария и три — летом. При этом в циклонических тропических и субполярных круговоротах (в них вода перемещается против часовой стрелки в северном полушарии и по стрелке — в южном) образуются зоны расхождения (дивергенции) вод, в которых развиваются восходящие токи воды, а в антициклонических субтропических круговоротах (направление вращения в них обратное но сравнению с циклоническими) поверхностные воды сходятся в центральных областях и погружаются там на глубину. Эти круговороты образуют гомологические пары, симметрично расположенные по отношению к экватору. Правда, эта симметрия (как и у большинства природных объектов) не обладает совершенством и полнотой, т. е. не идеальна и не касается всех признаков. Границы между круговоротами в открытом океане идут примерно в широтном направлении. Близ берегов зональное течение, общее для двух соседних круговоротов, раздваивается и между обеими крупномасштабными циркуляциями и берегом остаются треугольники не захватываемой ими воды — так называемые нейтральные области. В них существуют локальные круговороты меньшего масштаба.
В пределах каждого крупномасштабного круговорота набор водных масс по вертикали специфичен, а это означает, что в каждом круговороте имеется своя структура вод. Принято различать поверхностную, подповерхностную, промежуточную, глубинную и придонную водные массы, которые разделены слоями воды с повышенными градиентами свойств (в пределах каждой массы свойства не меняются или меняются очень постепенно). Выделяют два главных типа гидрологических структур — тропические, расположенные в тропиках и субтропиках, и субполярные, расположенные в умеренных широтах.
Для тропических структур характерны следующие водные массы и слои раздела между ними. Сверху находится поверхностный однородный (изотермический) слой, который характеризуется высокой температурой (20—30°), мало меняющейся по глубине. Толщина этого слоя в среднем равна 50—100 м (в центральных частях субтропических круговоротов до 200—250 м). Под изотермическим слоем температура воды сравнительно резко надает, и ее градиент до глубины 400—500 м имеет порядок 0,1 град/м. Одновременно резко возрастает плотность воды. Под слоем повышенного градиента температура продолжает понижаться, хотя и более медленно. До глубины около 1000—1500 м (в зависимости от структуры) это понижение происходит, однако, быстрее, чем на еще больших глубинах. Этот слой называют «главным термоклином», и его нижняя граница выражена не очень четко. Глубже него вода отличается низкой температурой (около 2°), мало меняющейся с глубиной. Это так называемые «глубинные однородные воды».
Резкое расслоение толщи воды по температуре, а следовательно, и но плотности чрезвычайно характерно для тропиков. Плотность воды зависит, однако, и от солености, а в большей части тропической зоны выражен промежуточный слой повышенной солености на глубине нескольких сот метров, т. е. в верхней части главного термоклина. В результате главный термоклин распадается на два слоя слой максимума солености до глубин 200—400 м и слой менаду ним и границей глубинных однородных вод, обозначаемый обычно как промежуточный минимум солености.
В субполярных водах весь столб воды сравнительно однороден по температуре. Главного термоклина там практически нет, и набор водных масс существенно отличается от характерного для тропической зоны. Поверхностный изотермический слой может нагреваться летом выше 10° (при толщине около 50 м) и охлаждаться зимой до 0—5° (иногда и до отрицательных значений температуры). Под этим слоем летом может существовать промежуточный холодный слой, в котором температура удерживается на уровне ее минимального зимнего значения; он может достигать глубины около 300 м. Промежуточный теплый слой с температурой на 1—2° выше, чем в предыдущем, располагается примерно до глубины 2000 м. Глубинные однородные воды мало отличаются по своим характеристикам от глубинных вод в тропических структурах.
Рассматривая вертикальную термическую структуру Мирового океана, нередко придают особое значение противопоставлению теплого поверхностного и холодного глубинного объемов водной толщи, называемых соответственно «термосферой» и «психросферой». Автор этих терминов — датский океанолог и ихтиолог А. Бруун принимал за границу таких слоев изотермическую поверхность 10°. Таким образом, в высоких широтах термосфера, естественно, отсутствует, в умеренных широтах она имеет сезонный характер, а в тропической зоне постояпна, но варьирует по толщине от 100 м в восточных районах океанов, характеризующихся подъемом глубинных вод, до 700 м в западных.
Каждый участок дна омывается водами того или иного слоя водной толщи, и поэтому дно по вертикали может подразделяться на те же участки, что и пелагиаль (тонкий слой воды у самого дна изучен недостаточно, и о его характеристиках судят в основном по характеристикам основного объема соответствующей водной массы). На дне океана выделяются, кроме того, участки (зоны), различающиеся режимом и скоростью осадконакопления, размерами частиц грунта и содержанием в нем органических веществ.
Почти все органическое вещество в океане первично продуцируется в приповерхностной зоне — в освещенных (эвфотических) слоях, где возможен фотосинтез, а затем разными путями проникает вглубь и служит той пищей, которая обеспечивает существование населения в лишенных света слоях воды и на дне. Роль органического вещества, создаваемого в лишенных света глубинах океана за счет первичных хемосинтетических процессов, невелика: оно составляет лишь доли процента от синтезируемого растениями.
Продукция фитопланктона определяется целым рядом факторов — интенсивностью солнечной радиации, содержанием биогенных солей (фосфатов, нитратов, силикатов) , толщиной верхнего однородного слоя, прозрачностью воды и т. д. Главным лимитирующим показателем практически везде служит все же концентрация в эвфотической зоне биогенных солей, основной запас которых находится в более глубоких слоях воды. Распределение этих солей в открытом океане обусловлено в первую очередь динамическими факторами, вызывающими подъем глубинных вод в верхний слой. В соответствии с этим районы наибольшей первичной продукции расположены в умеренных широтах, где обогащение поверхностных слоев питательными веществами происходит в результате зимнего конвективного перемешивания, а также в некоторых районах тропической области, характеризующихся наиболее интенсивным подъемом «свежих» глубинных вод. К ним относятся районы поднятия вод (апвеллингов) в зонах действия сгонных ветров у берегов, дивергенции течений (особенно субэкваториальная дивергенция) и центральные участки некоторых локальных круговоротов. Наименьшими показателями продукции фитопланктона отличаются обширные по площади области опускания вод в антициклонических субтропических круговоротах. Ежегодная суммарная величина первичной продукции составляет, по современным представлениям, не менее 60 млрд т углерода.
Продукция фитопланктона, представляющего собой самый низший трофический уровень в пищевых взаимосвязях населения пелагиали, служит основой общей биологической продуктивности океана и в конечном счета его промысловой продуктивности. Дальнейшее перераспределение органического вещества, синтезированного водорослями-продуцентами, осуществляется животными консументами — растительноядным и хищным зоопланктоном, а затем нектоном — и проходит, таким образом, через несколько трофических уровней. При переходе на каждый последующий уровень расходуется большая часть энергии (такие потери часто достигают 90%), и это ограничивает число уровней: в холодных водах их 3—5, в тропиках — до 7. В одних пелагических сообществах (их называют сбалансированными по трофическим циклам) продукция, ежегодно создаваемая на каждом уровне, нацело выедается хищниками следующего уровня, в других (несбалансированных) эта продукция длительно и существенно превышает выедание и ее избыток выносится течениями в другие участки или попадает на дно. Наименьшей сбалансированностью отличаются неритические сообщества, а максимальное приближение к сбалансированности имеет место в тропической части открытого океана, где наблюдается высокая стабильность в количественном соотношении организмов, стоящих на разных трофических уровнях.
Количественное распределение и продукция зоопланктона в океане в общих чертах соответствуют обилию растительной жизни. Наибольшая биомасса среднеразмерного зоопланктона («сетного» зоопланктона, или мезопланктона) наблюдается летом в умеренных и высоких широтах (прежде всего во фронтальных зонах), а также в экваториальных районах и районах глобальных подъемов вод в восточных частях океанов, наименьшая биомасса — в зонах антициклонических субтропических круговоротов, так называемых «океанических пустынях». Наибольшая продукция мезопланктона также создается в субполярных водах обоих полушарий и в водах прибрежных и экваториальных апвеллингов, а иногда и в районах локальных продуктивных пятен.
Распределение биомассы океанического макропланктона и нектона тоже подчинено широтной и циркумконтинентальной зональности. Прослеживаются три полосы повышенной биомассы — в северных умеренных, экваториальных и южных умеренных широтах, причем в экваториальной зоне количественные показатели обилия снимаются в направлении с востока на запад в Тихом и Дтлантическом океанах и с северо-запада на восток — в Индийском океане. Эти полосы перемежаются четырьмя полосами пониженной биомассы — арктической, двумя центральными (субтропическими) и антарктической, в которой биомасса все же значительно выше, чем в трех других. Во всех океанах значительная биомасса консументов высших трофических уровней (главным образом рыб) приурочена к нерито-океанической зоне, переходной между прибрежными и океанскими районами. В этих участках максимальная биомасса нектона наблюдается на западных окраинах океанов в зоне полярных фронтов и на их восточной периферии мористее районов пассатных апвеллингов и вдоль прилегающих фронтальных разделов. В Индийском океане, где северного полярного фронта и северо-восточного апвеллинга нет, повышенная биомасса наблюдается мористее районов Аравийского и Сомалийского сезонных апвеллингов. В экваториальных широтах количество макропланктона и нектона особенно велико на востоке океанов (продуктивные зоны Гвинейского и Панамского заливов и прилегающих вод) и в районах локальных апвеллингов Карибского моря — Мексиканского залива и австрало-азиатских морей.
Все перечисленные выше (а также некоторые другие) особенности открытого океана оказывают непосредственное влияние на пространственное распределение обитающих в его водах организмов, в том числе и рыб. Участки, различающиеся по своему населению, могут иметь границы, совпадающие с изобатами, изотермами, изогалинами, линиями равного содержания растворенного кислорода, первичной продукции, биомассы мезоплаиктона и т. д., однако чаще всего эти границы соответствуют разделам дискретных водных масс, на которые подразделяется водная толща. Поэтому многие исследователи считают, что деление океана на крупномасштабные биотопы — участки, имеющие однородное население, — в первую очередь определяется именно расчлененностью его объема (так называемая «гипотеза водных масс»).
Границы водных масс в придонных горизонтах неплохо совпадают с основными формами рельефа дна. В вертикальном плане океан подразделяется поэтому на следующие зоны: верхнюю (от поверхности до глубины 100— 200 м), среднюю (до глубины 1000—1500 м), нижнюю (до глубины 3000 м), глубинную (до глубины 6000 м) и сверхглубинную (рис. 1). В классификации участков океанической среды обитания наибольшее значение имеет их разделение на пелагические биотопы, с одной стороны, и донно-придонные — с другой. В пелагиали верхняя зона соответствует поверхностным и подповерхностным водам, средняя — промежуточным, нижняя — глубинным глубинная — придонным, а сверхглубинная — водам глубоководных желобов; на дне (в бентали) верхняя зона более или менее совпадает с шельфом; средпяя и нижняя — с материковым склоном, глубинная — с ложем океана, а сверхглубинная — с дном желобов. При этом нужно иметь в виду, что приведенные примерный глубины не могут рассматриваться как абсолютные показатели границ вертикальных зон: их положение сильна варьирует в разных районах Мирового океана.
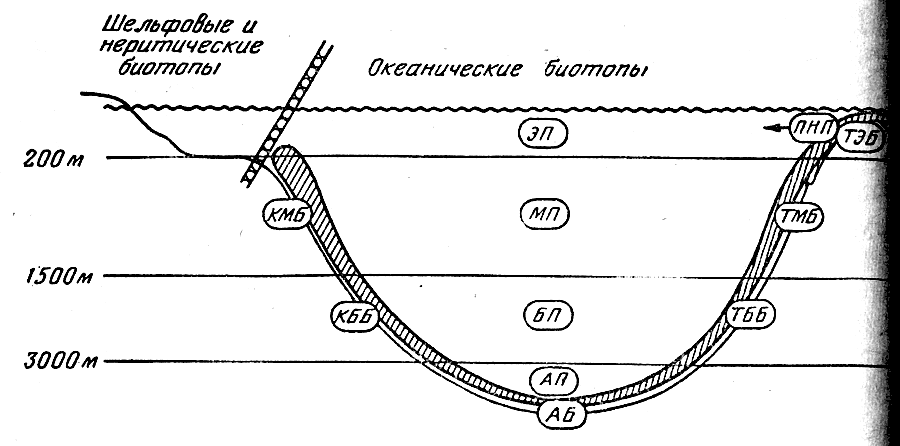
Рис. 1 . Вертикальная зональность океанических биотопов.
Названные вертикальные зоны в пелагиали вполне традиционно именуются эпи-, мезо-, бати-, абиссо- и хадопелагиалыо. Для бентали этих же зон в СССР обычна применяют названия сублитораль (на шельфе), «переходный горизонт» и батиаль (соответственно на верхней и нижней частях склона), абиссаль (на ложе океана) и ультраабиссаль (на дне глубоководных желобов). Им экбивалентны вертикальные зоны, названные Т. С. Рассом эпи-, мезо-, бати-, абиссо- и хадобенталью, и эта унифицированная терминология представляется более удобном для сопоставления биотопов дна и толщи воды (термин «батиаль» сохраняется при этом для обозначения всего биотопа материкового склона, т. е. идентичен понятию «мезобатибенталь»).
Население пелагиали контактирует с населением бентали во всех вертикальных зонах в придонных слоях водвой толщи, которые могут рассматриваться как биотоп вторичного сообщества — бентопелагического экотона (по определению К. В. Беклемишева, «экотон есть биотоп сообщества, состоящего из смеси видов двух соприкасающихся биоценозов»). В составе этого экотона в целом преобладают пелагические виды, а настоящие бентосные, как правило, находятся лишь па ранних стадиях жизненного цикла. В бентопелагиали имеются и специфические виды, и именно они составляют наиболее характерную часть ее населения. Это сообщество особенно четко обособлено в самой верхней вертикальной зоне (хорошо известное население неритической эпипелагиали), но оно существует во всем диапазоне океанских глубин. Бентопелагиаль, как и пелагиаль, является трехмерным биотопом и отличается в этом отношении от «плоской» двухмерной бентали, но объем придонной воды составляет ничтожную часть общего объема водной толщи, и в планетарном масштабе донный и придонный биотопы представляются тесно сопряженными (действительно, они не могут существовать в разделенном виде).
В дополнение к приведенной выше схеме вертикальной зональности А. П. Андрияшев выделил переуглубленные участки дна на шельфе — псевдобатиаль и изолированные от океанских глубин глубокие (более 3000 м) бассейны — псевдоабиссаль. Примеры псевдобатиального биотопа дают многочисленные депрессии на шельфе Антарктиды и в бассейне Северного Ледовитого океана, отделенные мелководьями от материкового склона; примерами псевдоабиссали (псевдоабиссобентали) являются Японское и Средиземное моря, соединяющиеся с океаном неглубокими проливами, обособленные котловины Полярного бассейна и т. д. Бесшельфовые подводные поднятия с глубинами порядка батиальных были выделены в талассобатиаль, и этот термин (также предложенный А. П. Андрияшевым) широко применяется в отечественной литературе. По аналогии с ним вершины подводных гор (часто их неправильно называют банками) с глубиной порядка 200 м и менее и «шельфы» малых островов были названы мной талассосублиторалью, или талассоэпибенталью. Кроме того, над высокими подводными поднятиями и около них (а иногда, по-видимому, и в других участках со слабо сбалансированными трофическими цикла ми) могут возникать условия, в некоторой степени ими тирующие особенности неритической зоны, и это позволяв ет обозначать такие участки как псевдонеритические (или талассоэпипелагические). Таким образом, в бентали и бентопелагиали Мирового океана три зоны — верхняя средняя и нижняя — существуют в двух модификациях субконтинентальной и талассной, а зона верхней пелагиали даже в трех — субконтинентальной (неритической) собственно океанической и талассной (псевдонеритической). Из семи субконтинентальных биотопов к океаническим относятся, однако, только донные и придонный биотопы материкового склона (соответствующие модифинации трех биотопов верхней зоны являются шельфо-нeритическими) (табл.1).
Эти наиболее крупномасштабные биотопы вертикального плана, как уже говорилось, делятся на биотопы подчиненных рангов, причем степень их возможного дробления в общем уменьшается от верхних слоев к нижним и от континентов к открытым водам. Из-за большего разнообразия условий в бентали степень дробности ее биотопов значительно больше, чем пелагических, которые как принято считать, представляют собой участки среды характеризующиеся наличием циркуляции, устойчивых в пространстве и времени.
Итак, система биотопических комплексов открытого океана имеет закономерную структуру и состоит из гомологичных частей. Примерами гомологичных биотопов в пелагиали могут служить парные крупномасштабным круговороты и их водные массы, расположенные симметрично по отношению к экватору в том же океане или занимающие сходное положение в других океанах (например, все субполярные или все центральные круговороты). В вертикальном расслоении водной толщи полная биотопическая гомология прослеживается только в пределах однотипных гидрологических структур — тропических и субполярных; так как если глубинные однородные по температуре воды (батипелагиаль) в обоих случаях вполне гомологичны, то поверхностный изотермический слоя в умеренных и высоких широтах в отличие от тропических имеет лишь сезонный характер. Поэтому нижняя граница эпипелагиали в общем совпадает с верхней границей скачка плотности воды (пикноклина), обусловленного в тропических водах понижением температуры, а в субполярных — увеличением солености. Мезопелагиаль в тропической зоне занимает слой основного термоклина — промежуточные воды с большими температурными градиентами, а за пределами тропиков — промежуточные холодные и теплые слои. Таким образом, тропическая и внетропическая мезопелагиаль сильно разнятся по условиям существования и не гомологичны между собой. Донные биотопы гомологичны между собой в той мере, в какой гомологичны водные массы, омывающие соответствующие участки дна. В пределах этих участков более дробные гомологи определяются типом осадконакопления.
Таблица 1
| вертикальные зоны и их примерные границы по глубине | модификации биотопов | ||
| океаническая | талласная | субконтинентальная | |
| верхняя (от поверхности до 100-200м) | эпипелагиаль | эпипелагиаль, эпибенталь, эпибентопелагиаль | все биотопы заняты шельфо-неритическими сообществами |
| средняя (от 100-200 до 1000-2000 м) | мезопелагиаль | мезопелагиаль, мезобентопелагиаль | мезопелагиаль, мезобентопелагиаль |
| нижняя от 1000-1500 до 3000 м | батипелагиаль | батибенталь, батибетопелагиаль | батибенталь, батибетопелагиаль |
| глубинная(глубже 3000м) | абиссопелагиаль, абиссобенталь, абиссобентопелагиаль | не существует | не существует |
Сообщества, населяющие биотопы открытого океана, исключительно разнообразны в видовом отношении, но в этой книге они рассматриваются на примере только одного их фрагмента — таксоцена рыб, или ихтиоцена. Таксоценом называют совокупность всех видов некоторой таксономической группы любой надвидовой категория входящих в состав одного биоценоза любого иерархического ранга. Необходимость в таком термине определя ется тем обстоятельством, что изучение конкретных сообществ крайне редко проводится по полному состав слагающих его видов. В этом, действительно, не всегда есть нужда, так как многие особенности биоценозов могут быть исследованы и на примере отдельных таксоценов. В отечественную ихтиологическую литературу понитие «ихтиоцен» было впервые введено именно для обозначения совокупности рыб как части единого сообщества, объединяемого общим биотопом. Впоследствии в этом же биоценотическом смысле употреблялись такие термины, как «ихтиоценоз», «рыбная часть сообщества»; «рыбное сообщество», «рыбное население» и «ихтиокомплеке», из которых первые четыре должны рассматриваться как полные синонимы «ихтиоцена», а последний лишен строго детерминированного смысла. Некоторое представление о составе и разнообразии ихтиоценов открытого океана дает рис. 2.
Основные океанические ихтиоцены — пелагический на бентический (донный) — традиционно рассматриваются в качестве самостоятельных категорий. Этого нельзя сказать о бептопелагическом (придонно-наддонном) ихтиоцене, реальность существования которого над материковым склоном и ложем океана часто недооценивается, несмотря на его полную гомологию сообществу рыб неритической эпипелагиали.
Бентопелагические рыбы образуют довольно разнородную ассоциацию, в которой представлены виды разного происхождения и разной экологии. Среди них могут быть выделены 1) рыбы, обитающие в узком (толщиной в несколько метров) слое над дном, 2) рыбы, живущие в толще вод над дном, но не только вблизи дна, но и в зная чительном (на десятки — сотни метров) удалении 3) рыбы, мигрирующие ночью от придонных горизонтов в верхние слои водной толщи (эта группировка существует только в среднеглубинной зоне). Все бентопелагические рыбы почти не имеют прямого контакта с грунтом (некоторые, правда, откладывают донную икру или временами поедают донных животных) и этим отличаются от бентических рыб, которые могут лежать на дне, oпираться на него плавниками или ползать по нему, хотя и не теряют способности к плаванию. Для обозначения всей совокупности донно-придонно-наддонных рыб (при противопоставлении их чисто пелагическим видам) можно предположить название «бентопелагический комплекс».
По типу связи со средой обитания все океанические организмы группируются в три основных категории — планктон, нектон и бентос, причем рыбы представлены в каждой из этих жизненных форм. Планктон парит в толще воды и, не имея возможности длительно поддерживать направленную горизонтальную скорость, соизмеримую со скоростью перемещающихся вод, проявляет способность к активным миграциям только в вертикальном направлении. Нектон объединяет хорошо плавающих животных, «свободных в выборе своего пути» (по выражению Э. Геккеля) и произвольно совершающих не только вертикальные, по и горизонтальные перемещения, которые иногда достигают очень большой протяженности. Живущие на дне организмы — бентос — подразделяются па две группы: одну образуют сессильные формы (сидячие или малоподвижные), другую — вагильные, или мигрантные (способные к перемещениям по дну). Конечно, типичные представители всех трех основных категорий связаны многочисленными переходами: многие рыбы принадлежат, например, к нектопланктону и нектобентосу. Кроме того, многие животные бентоса и почти все нектонные животные проходят в своем развитии планктонную стадию. Плавучие икринки и личинки костистых рыб объединяются, например, под собирательным названием «ихтиопланктон».
Ихтиопланктон открытого океана включает не только ранние стадии развития океанических рыб (естественно, исключая щележаберных — акулы и скаты с самого начала жизни принадлежат к нектону), но и личинок и мальков множества шельфо-неритических видов, которые попадают в эпипелагиаль в результате пассивного выноса. В тропических водах его характеризует эфемерность существования — ведь развитие икры продолжается, как правило, не более 2 суток (правда, у макрелещуковых и летучих рыб — 2 недели, у вогмеровых рыб — до 2 месяцев), а личинки находятся в составе планктона в среднем около 8 недель. В более холодноводных районах эти сроки удлиняются, и длительность предмалькового периода жизни достигает у большинства видов 4—6 месяцев. Сильно растянуто личиночное развитие и у некоторых тепловодных форм, в частности у многих камбаловых и угреобразных рыб (личинки европейского угря, например, проводят в ихтиопланктоне до 3 лет), и это сильно способствует их дальнему разносу и широкому pacnpoстранению.
Икринки и личинки имеют приспособления к планктонному существованию, способствующие парению в толще воды: разного рода выросты и придатки на теле, удлиненные лучи плавников, а также жировые включения и газовые пузырьки (рисунок 3). Как правило, они сильно отличаются по всему своему облику от взрослых особей и в ряде групп претерпевают резкий метаморфоз (изменение строения) в ходе индивидуального развития. Поэтому установить принадлежность раннего малька, а тем более личинки (не говоря уже об икринке) к тому или иному виду, роду и даже семейству во многих случаях совсем непросто.
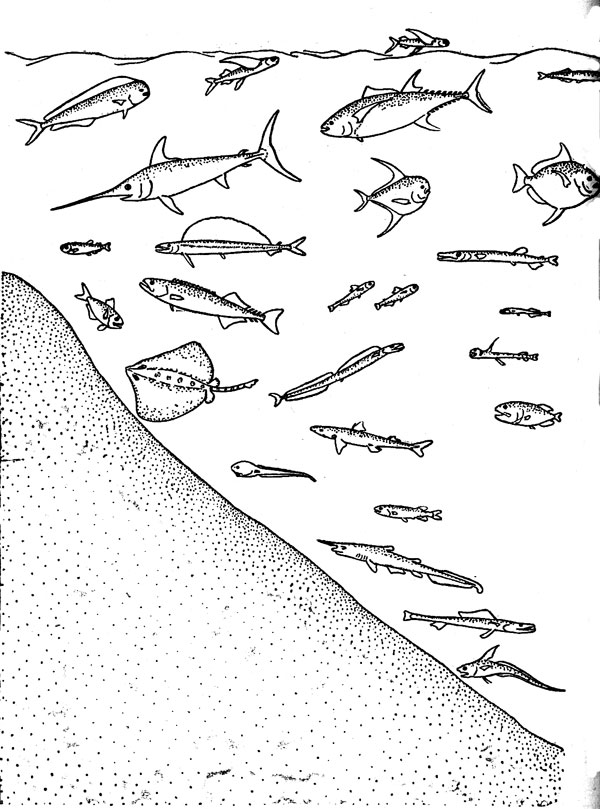
Рис. 2. Характерные рыбы океанических ихтиоценов (схема)

Рис. 2. Характерные рыбы океанических ихтиоценов (схема)
Особенно своеобразны прозрачные листовидные личинки угрей и близких к ним групп рыб. В прошлом столетии этих «рыбок» относили к особому роду Leptocephalus, и это название конечно, не в качестве родового осталось за ними до сих пор. Лептоцефалы некоторых видов достигают очень солидных размеров - рекорд (184 см) принадлежит L. giganteus (эта личинка относится к отряду спиношипообразных). Чтобы вырасти до такого размера, нужно, казалось бы, неплохо подкормиться, но, как ни странно, в кишечниках у этого гиганта или каких-либо других лептоцефалов никогда не находили даже следов пищи. Предполагают поэтому, что они могут усваивать растворенное в воде органическое вещество непосредственно через эпидермис. Согласно другому предположению, лептоцефалы высасывают протоплазматические соки из планктонных организмов, протыкая их покровы острыми зубами.
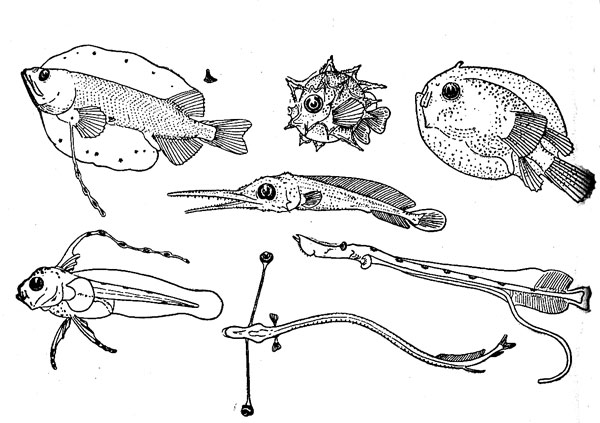
Рис. 3. Личинки океанических рыб
...В одном из первых тропических плаваний «Витязя» по судовой трансляции как-то прозвучало сенсационное объявление: «Сотрудниками ихтиологического отряда только что впервые в практике советских морских экспедиций выловлен экземпляр меч-рыбы. Сейчас рыба находится в лаборатории, где ее может увидеть каждый желающий». Через несколько минут помещение маленькой ихтиологической лаборатории было забито возбужденным народом, а толпа опоздавших теснилась у двери и иллюминаторов, выходящих на палубу. Они считали себя обманутыми, так как обещанной огромной рыбины, вооруженной мощным мечом, нигде не было видно. И тем не менее в объявлении не было неправды: в плоской чашке на столике бинокулярного микроскопа лежала полосатая игловидная личинка, усеянная мелкими шипиками и не превышавшая полутора сантиметров в длину... Через несколько лет при ином стечении жизненных обстоятельств эта моч-рыба не обманула бы ничьих ожиданий...
Мне никогда не приходилось всерьез заниматься изучением пхтиопланктопа (для этой исключительно тонкой работы нужны особые качества, которыми я не обладаю), хотя не раз доводилось иметь с ним дело, особенно в первых экспедициях — в те давно прошедшие времена одна из моих лаборантских обязанностей состояла в отборе икры и личинок рыб из планктонных проб. Эта нудная, как тогда казалось, работа (она и вправду очень однообразна) дала мне, однако, возможность воочию познакомиться с изумительным многообразием строения рыб на ранних стадиях развития, когда они так разительно непохожи на своих родителей. Я не могу забыть изумления и восхищения, которые мне доставляли тогда первые встречи с личинками листовидных совершенно прозрачных угрей, шарообразных колючих рыб-лун, стебельчато-глазых идиакантов, исключительно разнообразных по форме тела миктофид (взрослея, они становятся гораздо более схожими между собой), камбал, тунцов, рыб-топориков и прочими удивительными существами, которых так занимательно разглядывать под увеличением. С тех пор я сохраняю интерес к ихтиопланктону (мне даже удалось впервые определить видовую принадлежность нескольких неописанных личинок). Как и многие другие ихтиологи, я твердо убежден, что признаки ранних стадий рыб должны обязательно приниматься в расчет при изучении их систематики и филогении.
Итак, реальность существования и относительная обособленность планктона и нектона, несмотря на их не всегда четкое разграничение, не вызывают сомнений. Эти группировки представляют собой две крупные жизненные формы пелагического населения — пассивную и активную, из которых каждая по-своему хорошо приспособлена к обитанию в толще воды. Ю. Г. Алеев предложил использовать для разделения планктона и нектона безразмерный физический показатель — соотношение инерционных эффектов и вязкости, характеризующее процесс движения и называемое числом Рейнольдса (Re). Плавание планктона, по его мнению, происходит в режиме Re<2,0*10M>7 (как правило, Re<5,0*10>5), а нектона в режиме Re>5,0*103 (как правило, Re>105), причем первый случай как бы соответствует приспособлению к существованию в условиях ламинарных токов воды, а второй — в условиях турбулентных течений.
Форма тела нектонных рыб соответствует их активному образу жизни. Наиболее быстрым и подвижным хищникам — тунцам, акулам и др. — свойственна возникшая конвергентным путем торпедообразная форма тела. У некоторых рыб (меч-рыба, марлины) оно снабжено; спереди еще особым рострумом-обтекателем, который, как предполагает В. В. Овчинников, турбулизирует набегающий поток и, подобно выступу в подводной части носа современных быстроходных судов, сильно уменьшает лобовое сопротивление. Важную роль в обеспечении возможности быстрого движения имеют и хвостовые кили, увеличивающие поперечную жесткость хвостового стебля и выполняющие роль горизонтальных стабилизаторов. Для нектоирых рыб довольно обычно также симметрично сжатое с боков тело, в той или иной степени вытянутое в длину, или стреловидное тело с отставленными кзади спинным и анальным плавниками.
В открытом океане минимальные размеры нектонных рыб могут, по-видимому, находиться в пределах от 15 до 20—30см Еще раз повторю, что разграничение планктона и нектона остается очень условпым. Я думаю, что ни размеры, ни крейсерские скорости, ни числа Рейнольдса сами по себе недостаточны для разграничения этих жизненных форм. Нужно учитывать пространственный масштаб биотопа — нектон небольшого озера и нектон открытого океана не могут, по моему мнению, иметь одинаковые характеристики..
Планктонные рыбы обладают весьма ограниченными; возможностями для активного плавания на всех стадиях жизненного цикла. К этой группе принадлежат не только карликовые (длина 2—5 см) рыбки, населяющие глубокие слои пелагиали — циклотоны и другие представите ли семейства гоностомовых (Gonostomatidae), меламфайя мелкие светящиеся анчоусы и т. д., но и мпогие более крупные (длина до 30—50 см) малоактивные обитателя этого биотопа — нитехвостые угри, мешкороты, хаулиоды и др. К планктону (частично к нектопланктону) относятся и все рыбы, движущиеся при помощи ундулирующих, (колебательных) движений плавников — удильщики, рыбы-луны, сельдяные короли. А ведь некоторые из них; достигают очень внушительных размеров — нескольким метров. Такие рыбы почти не способны противостоять встречному движению воды, и это делает их объектом пассивного горизонтального переноса течениями.
В пространственном распределении всех организмов планктона (в том числе, естественно, и планктонных рыб) решающую роль играет пассивный перенос течениями. Нектонные животные активно перемещаются в продолжение своего жизненного цикла, мигрируя в пределах районов, характеризующихся необходимыми для вида в тот или иной период условиями существования. Это обстоятельство приводит к довольно важным различиям в функциональной структуре ареалов планктонных и нектонных рыб.
В ареалах планктона выделяются следующие структурные части: 1) основа ареала, ограниченная, как принято считать, замкнутым круговоротом ( Существование таких круговоротов не всегда представляется обязательным, так как сохранение планктонной популяции в каком-либо участке области обитания в принципе может быть обеспечено и за счет переноса в системе вихревого движения; существенно в этом случае лишь соблюдение одного условия: экспатриация (вынос) из основы ареала и смертность не должны превышать пополнения популяции за счет размножения особей.), в котором может существовать независимая популяция вида; 2) нестерильная область выселения, приуроченная к выносящим течениям и населенная зависимыми, но самовоспроизводящимися популяциями, непрерывно пополняемыми из основы ареала; 3) стерильная область выселения — участки, пригодные для обитания взрослых рыб, но не пригодные для размножения, где происходит постепенное вымирание зависимых популяций. Основой ареалов планктонных видов могут служить как крупномасштабные океанические циркуляции, так и круговороты, расположенные в областях схождения или расхождения течений у берегов. На этом основании К. В. Беклемишев разделил планктонные виды на «океанические» и «дальненеритические», причем последние (в виде зависимых популяций) могут далеко распространяться в открытые воды с течениями, идущими от берегов. У многих широко распространенных организмов основа ареала может занимать сразу несколько круговоротов.
Константин Владимирович Беклемишев (1928-1983) внес значитальный вклад в разработку современных концепций пелагической биогеографии. Его фундаментальная работа «Экология и биогеография пелагиали», защищенная в 1967 г. в качестве докторской диссертации и увидевшая свет в 1969 г. в виде монографии, сыграла очень большую роль в становлении отечественной школы морских биогеографов.
Я был знаком с Константином с детских лет, как говорится, по семейной линии — его мать Нина Петровна и мой отец были одвокурсниками Пермского университета и вместе слушали лекции его отца - одного из крупнейших отечественных зоологов Владимира Николаевича Беклемишева (с 1934 г. он был профессором МГУ) В конце 40-х годов я часто бывал в этой прекрасной семье, сохравившей лучшие традиции русской интеллигенции. В субботние (а может быть, воскресные) вечера в доме собиралось много молодежи - студентов и школьников, при активном участии взрослих за чайным столом обсуждались самые разные темы, инсцениривались шарады — все чувствовали себя просто и непринужденво, но в то же время совсем незаметно впитывали новые знания в представления. Неудивительно поэтому, что Костя был человеком высочайшей культуры — он свободно владел французским, английским и немецким языками, прекрасно ориентировался в, гуманитарных науках и, конечно, имел глубочайшие познания в естественных, прежде всего в различных областях биологии. Совместная работа с Константином Беклемишевым (мы опубликовали в соавторстве 6 статей) дала мне очень много и сильно способствовала формированию моих научных взглядов.
Ареалы нектона представляют собой районы, границы которых определяются активными перемещениями caмих рыб, не выходящих за пределы вод с благоприятными для них условиями. В то же время способность к продолжительному плаванию дает этим рыбам возможность дифференцированно использовать разные части ареала в продолжение жизненного цикла. В соответствии с этим в ареалах нектонных рыб различают 1) область размножения (нереста), или репродуктивную часть ареала; 2) нагульную, или вегетативную, часть ареала, используемую видом для откорма путем двусторонних активных (иногда — за счет разноса молоди — пассивных в начальной фазе) миграций, нередко имеющих сезонный характер; 3) область пассивного выселения планктонных икринок, личинок и мальков. Структура ареала нектобентических рыб в принципе та же, но их миграции не имеют значительной протяженности.
Различия в структуре ареалов планктона, нектопа и бентоса не препятствуют проведению единого биогеографического деления Мирового океана, хотя до самого по следнего времени такая возможность представлялась сомнительной. Традиционно (еще со времен одного из основоположников морской зоогеографии А. Ортмаппа) анализ географического распространения организмов производился раздельно для толщи вод и для дна, а также для главных вертикальных зон. Такое разделение обусловлено резкими различиями в распространении населения толщи воды и дна, имеющими в своей основе кардинальные различия подвижных и неподвижных блотопов. В то же время известно, что основные биогеографические границы достаточно четко связаны с гидрологической структурой океана и совпадают с ее главными элементами — гидрологическими фронтами, зонами конвергенции и дивергенций течений. Поэтому широтная зональность в распределении пелагических и донных организмов сохраняется от поверхности воды до ложа океана, хотя дробность возможного биогеографического деления всегда уменьшается с глубиной.
При самом крупномасштабном районировании Мирового океана, основанном главным образом на распространении крупных систематических групп (ранга семейства и выше) выделяют несколько областей, объединяемых в три циркумглобальных биогеографических царства, или надобласти, — Аркто-Бореальное, Тропическое (Тропическо-Субтропическое) и Нотально-Антарктическое. В этом делении отражено наиболее серьезное из существующих различий региональных флор и фаун — обусловленное историческими причинами противопоставление холодноводных и тепловодных сообществ.
Районирование верхних и средних слоев пелагиали в пределах надобластей проводится главным образом на основе распределения отдельных видов планктона и нектона. Повторяющиеся видовые ареалы могут быть сгруппированы по сходству формы и географического положения в несколько основных типов или «географических элементов» фауны, что позволяет выделять участки среды того или иного биогеографического ранга. Наибольшая дробность районирования выявляется при анализе распространения планктонных организмов.
В целом биогеографическое районирование океанской эпи- и мезопелагиали подчинено широтной зональности, нарушаемой в той или иной степени только в приконтинентальных участках. В Мировом океане достаточно четко различаются следующие широтные зоны: 1) арктическая и 2) бореальная (с высоко- и низкобореальной подзонами), входящие в состав Аркто-Бореальной надобласти; 3) северная субтропическая, 4) тропическая (с северной центральной, экваториальной и южной центральной подзонами) и 5) южная субтропическая в составе Тропическо-Субтропической надобласти; 6) потальная и 7) антарктнческая (с низко- и высокоантарктической подзонами) в составе Ноталыю-Антарктической надобласти. В Ипдийском океане трех северных зон нет, и счет начинается тропической зоны (точнее говоря, с ее экваториальной подзоны). Деление бореальной зоны на подзоны отчетливо выражено только в Тихом океане, тогда как в Атлантике они значительно менее обособлены. Расчленение тропической зоны на экваториальную и центральные подзоны заметно главным образом по распространению планктонных видов, но благодаря эффекту экспатриации (выселения) границы этих подзон, как правило, оказываются сильно размытыми. Тропическая и субтропические зоны (выделение последних основано главным образом на особенностях распространения нектонных рыб и кальмаров) также разделены переходными полосами, населенными смешанной фауной. Еще более широкие «переходные зоны» расположены на периферии Тропическо-Субтропической надобласти, т. е. у ее границ с бореальной и нотальной зонами. Особенно хорошо такой участок, называемый «переходной зоной Южной субтропической конвергенции», выражен в южном полушарии. В некоторых систематических группах (в частности, среди макропланктонных рыб) здесь наблюдается высокий видовой эндемизм (более 30%), но в других таксоценах пелагического сообщества он почти отсутствует.
Границы всех широтных зон и подзон достаточно четко выражены в средних частях океанов и еще более резки в районах схождения течений у берегов. К таковым относятся, в частности, фронтальные зоны между субполярными и центральными круговоротами в западных частях океанов, соответствующие северной и южной границам Тропическо-Субтропической надобласти. На востоке эти границы, напротив, находятся в зонах расхождения течений и представляют собой обширные участки переходного типа, расположенные в Канарском, Бенгельском, Калифорнийском и Перуанском течениях. Субтропический зоны и центральные подзоны также сильно перекрываются в восточных частях океанов.
Глядя с палубы на однообразную поверхность океана, трудно вообразить, что где-то под килем проходят в толще воды незримые, но реальные зоогеографические границы. Еще труднее представить себе, что эти границы вполне равнозначны тем, которые разделяют столь резко различающиеся ландшафты суши - тундру и тайгу, степь и пустыню. Между тем в каждом участке пелагиали есть характерные рыбы, которые с первого взгляда распознаются специалистами. Поэтому даже на коллег-океанологов, не связанных с биологией моря, производит большое впечатление, когда ихтиолог, лишь глянув на банку с уловом мезопслагических рыб, уверенно называет место, где была получена проба. Обратный вариант — предсказание состава улова, когда сеть еще находится в воде,— сложнее: он возможен только там, где ихтиофауна не отличается значительным разнообразием. Лет тридцать назад, участвуя в рейсе в северной части Тихого океана, я завоевал такими прогнозами большое уважение у сурового начальника экспедиции — известного специалиста по подводной технике Н. Н. Сысоева. В этом относительно бедном видами районе мне удавалось угадывать не только видовой состав улова на каждом горизонте траления, но и примерное число экземпляров всех видов. Все это я говорю отнюдь не для того, чтобы похвастаться какими-то необыкновенными способностями. Моя основная мысль такова: морская зоогеография — точная и строгая наука, и знание ее принципов очень помогает в самых разных ситуациях. Приведу хотя бы такой пример. Одна из сотрудниц нашего института, обрабатывая ихтиопланктонные пробы из Южно-Китайского моря, обнаружила в них личинок миктофид, которых она отнесла к виду, характерному для субарктической Пацифики. С зоогеографической точки зрения это гак же невероятно, как встреча с белым медведем в Сахаре, и, действительно, определение оказалось ошибочным.
Дальненеритические участки занимают большую площадь в восточных частях океанов. Особенно своеобразный регион образует восточная тропическая Пацифика, и пределах которой (между 20° с. ш. и 10° ю. ш.) находится не менее трех «нейтральных» областей. Фауна этого региона заметно обеднена за счет отсутствия многих широко распространенных тропических видов, но содержит значительное число эндемичных форм. Основываясь на данных о распространении мелких мезопелагических рыб, А. Эбелинг придавал этому участку Тихого океана такой же ранг «первичного зоогеографического региона», как и всей остальной части Тропическо-Субтропической иадобласти Мирового океана.
Схемы биогеографического районирования верхних и средних слоев пелагиали, предложенные разными авторами, при общем принципиальном сходстве заметна различаются в деталях. Основные особенности пространственного распределения планктона и нектона отражает, однако, следующая система ранжированных регионов, в основу которой положена рассмотренная выше зональность: Арктическая область с атлантической и тихоокеанской частями, Нереальные атлантическая и тихоокеанская области (с высокобореальными и низкобореальными провинциями); Атлантическая, Индо-Западно-тихоокеанская ((Индовестпацифическая) и Восточно-Тихоокеанская тропические области (две первых с северными субтропическими, экваториально-центральными и южными субтропическими провинциями); Нотальная область и Антарктическая область, в пределах которых различают (в значительной степени условно) атлантические, индоокеанские и тихоокеанские части.
В батипелагиали различия между тропическими и умеренно-высокоширотными ихтиофаунами также выражены вполне четко и полностью оправдывают выделение Тропическо-Субтропической, Аркто-Бореальной и Нотально-Антарктической надобластей, разделенных переходными зонами. Для деления этих надобластей на более дробные регионы данных пока недостает. Следует отметить, однако, что несмотря на большее однообразие условий среды, ареалы многих батипелагических рыб по форме и положению очень сходны с ареалами рыб мезопелагиали. Объяснение этому состоит, по всей видимости, в том, что раннее развитие всех таких рыб проходит в верхних слоях воды.
Изменения видового состава населения бентали также происходят на границах водных масс (эти изменения связаны с температурным режимом, типом донных отложепий, биологической продуктивностью водной толщи). Кроме того, в распространении донных и придонных животных (включая рыб) очень большое значение принадлежит так называемому генетическому (или историческому) фактору — положению того или иного участка относительно основных «эволюционных центров», отличающихся длительной стабильностью условий среды и действующих в качестве «генераторов» новых таксонов.
Не останавливаясь на деталях и частностях, нужно прежде всего отметить, что в бентали выделяются те же биогеографические области, что и в нелагиали. На шельфе, т. е. в эпибентали, эти области подразделяются на многочисленные провинции, которых только в Тихом океане насчитывается, к примеру, около 30. С увеличением глубины степень дробности районирования уменьшается, хотя в верхней части склона (мезобенталь) она еще не имеет отличий от самой верхней зоны. На дне океана (в абиссобентали) прослеживаются только сильно размытые границы зоогеографических областей.
Таким образом, открытый океан предоставляет своему населению (в том числе и рыбам) достаточно разнообразные условия жизни, сильно различающиеся по многим абиотическим и биотическим показателям. Естественно поэтому, сообщества и ихтиоцены разных структурных частей этого мегамасштабного биотопа имеют глубокие различия и достаточно четко обособлены по систематическому составу и по экологии входящих в них видов. Важнейшими подразделениями открытого океана, также имеющими глобальный масштаб, следует считать верхнюю пелагиаль (в пределах продуцирующего эвфотического слоя), глубоководную пелагиаль и совокупность донно-придонных участков. Обзору и характеристике соответствующих ихтиоценов посвящены три следующие главы.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://fish.geoman.ru/ 'Иллюстрированная энциклопедия рыб'